タイトル
2011年度 奨励賞 田中 謙二
概要

グリア研究展開に必要な技術の開発
Technological development toward progress in glial cell research
慶應義塾大学部医学部 精神神経科学教室 田中謙二
グリアとは何か、それを知るために神経化学会に入り、10年以上研究してきた。その10年の研究で分かったことが2つある。1つは、グリアの機能は他の細胞の機能変化を通して理解されることである。グリアと神経の相互作用、グリアと血管の相互作用など、常に何かのシステムの変化を通してグリアの機能は記載される。もう1つの分かったことは、グリア研究はin vivo研究を中心に展開していくということである。
前者の相互作用を明らかにする研究で大事なのは、起点を明確にすることである。起点が明確になれば、グリアから神経への一方向性の作用、神経からグリアへの一方向性の作用を独立に調べることが出来、相互作用の解析が深まる。
グリアの複雑な形態はin vivo研究でのみ観察可能で、そのダイナミズムはin vivo研究でのみ追究できる。構造は機能に裏打ちされているので、グリアの形を保ったままグリアの機能を調べるべきである。
細胞種特異的に、in vivoでグリアの機能を操作する技術は、両者を満たすうえで必須の技術になる。オプトジェネティクスは光で細胞の機能を操作する技術であるが、操作に使用するプローブ分子を細胞種特異的に発現させることが出来れば細胞種特異的な操作が可能になる。
筆者は光感受性タンパクであるチャネルロドプシン(ChR2)、アーチロドプシン(ArchT)をグリア細胞3種に発現させ、光でグリアの機能を操作する実験系を構築した。ChR2やArchTはイオンコンダクタンスがとても小さいために、膜電位を操作するには大量の発現を必要とする。
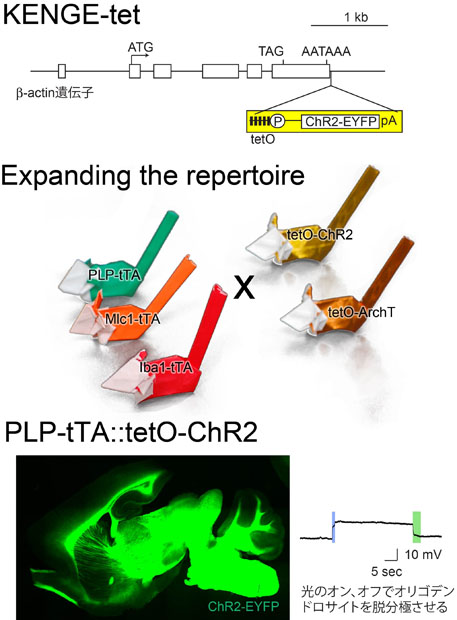
グリア細胞に大量に外来分子を発現させるために、テトラサイクリン遺伝子発現誘導システム(tetシステム)を利用した。Tetシステムは細胞種特異的発現を担保するtTAマウスと外来遺伝子の発現を増幅するtetOマウスとを交配させるシステムである。従来の方法で作成されるtetOマウスでは膜電位を操作するに足る発現量を確保することが難しかったが、tetOマウスを-actin遺伝子座にノックインすることで遺伝子発現が劇的に改善し(Knockin-mediated ENhanced Gene Expression-tet: KENGE-tet, 図の上段)、ChR2やArchTが機能するだけの十分な発現量を確保することが出来た[1]。
アストロサイト、オリゴデンドロサイト、ミクログリアだけにtTAを発現するマウス(Mlc1-tTA[2]、PLP-tTA[3]、Iba1-tTA[1])を筆者がすべて作成し、光操作が可能な組み合わせを一揃えすることが出来た(図の中段)。
オリゴデンドロサイトの光操作の一例を示す(図の下段)。PLP-tTAとtetO-ChR2のダブルトランスジェニックマウスでは、成熟オリゴデンドロサイトだけにChR2が発現する。光を照射すれば20 mV程度の脱分極がおこる。神経活動の亢進に伴って、近傍のオリゴデンドロサイトの膜電位が15-20 mV脱分極することが知られているので、そのオリゴニューロン相互作用を光で模倣できるのである。
このように、オプトジェネティクスを用いたグリア細胞の光操作によって、グリア細胞を起点とした状態変化を引き起こすことが出来る。他のシステムとの相互作用を解析するには優れた実験方法と思われる。また、遺伝子改変マウスを手に入れて、光を照射するだけでグリア細胞を操作できるので、グリア細胞の構造を保ったままグリア細胞の機能を調べることが出来る[4, 5]。
1. Tanaka, K.F., et al., Expanding the repertoire of optogenetically targeted cells with an enhanced gene expression system. Cell Rep, 2012. 2(2): p. 397-406.
2. Tanaka, K.F., et al., Flexible Accelerated STOP Tetracycline Operator-knockin (FAST): a versatile and efficient new gene modulating system. Biol Psychiatry, 2010. 67(8): p. 770-3.
3. Inamura, N., et al., Gene induction in mature oligodendrocytes with a PLP-tTA mouse line. Genesis, 2012. 50(5): p. 424-8.
4. Okada, Y., et al., Preinspiratory calcium rise in putative pre-Botzinger complex astrocytes. J Physiol, 2012. 590(Pt 19): p. 4933-44.
5. Sasaki, T., et al., Application of an optogenetic byway for perturbing neuronal activity via glial photostimulation. Proc Natl Acad Sci U S A, 2012. 109(50): p. 20720-5.
グリア研究展開に必要な技術の開発
Technological development toward progress in glial cell research
慶應義塾大学部医学部 精神神経科学教室 田中謙二
グリアとは何か、それを知るために神経化学会に入り、10年以上研究してきた。その10年の研究で分かったことが2つある。1つは、グリアの機能は他の細胞の機能変化を通して理解されることである。グリアと神経の相互作用、グリアと血管の相互作用など、常に何かのシステムの変化を通してグリアの機能は記載される。もう1つの分かったことは、グリア研究はin vivo研究を中心に展開していくということである。
前者の相互作用を明らかにする研究で大事なのは、起点を明確にすることである。起点が明確になれば、グリアから神経への一方向性の作用、神経からグリアへの一方向性の作用を独立に調べることが出来、相互作用の解析が深まる。
グリアの複雑な形態はin vivo研究でのみ観察可能で、そのダイナミズムはin vivo研究でのみ追究できる。構造は機能に裏打ちされているので、グリアの形を保ったままグリアの機能を調べるべきである。
細胞種特異的に、in vivoでグリアの機能を操作する技術は、両者を満たすうえで必須の技術になる。オプトジェネティクスは光で細胞の機能を操作する技術であるが、操作に使用するプローブ分子を細胞種特異的に発現させることが出来れば細胞種特異的な操作が可能になる。
筆者は光感受性タンパクであるチャネルロドプシン(ChR2)、アーチロドプシン(ArchT)をグリア細胞3種に発現させ、光でグリアの機能を操作する実験系を構築した。ChR2やArchTはイオンコンダクタンスがとても小さいために、膜電位を操作するには大量の発現を必要とする。
グリア細胞に大量に外来分子を発現させるために、テトラサイクリン遺伝子発現誘導システム(tetシステム)を利用した。Tetシステムは細胞種特異的発現を担保するtTAマウスと外来遺伝子の発現を増幅するtetOマウスとを交配させるシステムである。従来の方法で作成されるtetOマウスでは膜電位を操作するに足る発現量を確保することが難しかったが、tetOマウスを-actin遺伝子座にノックインすることで遺伝子発現が劇的に改善し(Knockin-mediated ENhanced Gene Expression-tet: KENGE-tet, 図の上段)、ChR2やArchTが機能するだけの十分な発現量を確保することが出来た[1]。
アストロサイト、オリゴデンドロサイト、ミクログリアだけにtTAを発現するマウス(Mlc1-tTA[2]、PLP-tTA[3]、Iba1-tTA[1])を筆者がすべて作成し、光操作が可能な組み合わせを一揃えすることが出来た(図の中段)。
オリゴデンドロサイトの光操作の一例を示す(図の下段)。PLP-tTAとtetO-ChR2のダブルトランスジェニックマウスでは、成熟オリゴデンドロサイトだけにChR2が発現する。光を照射すれば20 mV程度の脱分極がおこる。神経活動の亢進に伴って、近傍のオリゴデンドロサイトの膜電位が15-20 mV脱分極することが知られているので、そのオリゴニューロン相互作用を光で模倣できるのである。
このように、オプトジェネティクスを用いたグリア細胞の光操作によって、グリア細胞を起点とした状態変化を引き起こすことが出来る。他のシステムとの相互作用を解析するには優れた実験方法と思われる。また、遺伝子改変マウスを手に入れて、光を照射するだけでグリア細胞を操作できるので、グリア細胞の構造を保ったままグリア細胞の機能を調べることが出来る[4, 5]。
1. Tanaka, K.F., et al., Expanding the repertoire of optogenetically targeted cells with an enhanced gene expression system. Cell Rep, 2012. 2(2): p. 397-406.
2. Tanaka, K.F., et al., Flexible Accelerated STOP Tetracycline Operator-knockin (FAST): a versatile and efficient new gene modulating system. Biol Psychiatry, 2010. 67(8): p. 770-3.
3. Inamura, N., et al., Gene induction in mature oligodendrocytes with a PLP-tTA mouse line. Genesis, 2012. 50(5): p. 424-8.
4. Okada, Y., et al., Preinspiratory calcium rise in putative pre-Botzinger complex astrocytes. J Physiol, 2012. 590(Pt 19): p. 4933-44.
5. Sasaki, T., et al., Application of an optogenetic byway for perturbing neuronal activity via glial photostimulation. Proc Natl Acad Sci U S A, 2012. 109(50): p. 20720-5.