22. 記憶と関連するスパインを可視化・光操作する新技術の確立
東京大学・院・医・構造生理部門
白井福寿、林(高木)朗子
東京大学・院・医・構造生理部門
白井福寿、林(高木)朗子
Labelling and optical erasure of synaptic memory traces in the motor cortex. Akiko Hayashi-Takagi (CA), Sho Yagishita, Mayumi Nakamura, Fukutoshi Shirai, Yi Wu, Amanda L. Loshbaugh, Brian Kuhlman, Klaus M. Hahn, and Haruo Kasai (CA), Nature, 2015, Vol 525, 333-338.
DOI 10.11481/topics22
掲載日:2016年3月7日 登録日:2016年5月26日
はじめに
脳内の数百億もの神経細胞は、神経突起を介して互いに複雑に連結し合い、膨大な神経回路網を形成しています。これらの神経突起は直接連結しているのではなく、シナプスと呼ばれる数十nmの隙間があります。神経細胞体の興奮がシナプスまで到達すると、シナプス間隙より神経伝達物質が分泌され、放出された伝達物質は隣の神経細胞のシナプス後膜に結合します。このようなシナプス入力が積算されると神経細胞は興奮し、次々と隣の神経細胞に電気的信号を送ることができます。大脳皮質のシナプスの多くは、神経突起上にあるマッシュルームのような小突起(スパイン)の上に形成されます。スパインは、大きさとそのスパイン上に形成されるシナプスの伝達効率が非常に良く相関することが知られています1。重要なことは、スパインが学習・記憶により劇的に増大したり、さらには新しいスパインが構築され既存のスパインが排除されたりする点です。すなわち、スパインの形態変化により神経回路網は動的に変化するので、スパインは学習・記憶・認知の細胞基盤であると考えられてきました。しかし、動物の生きたままの脳内で、増大・新生したスパインを特異的に標識・操作する手法が無かったため、スパインと学習・記憶との因果関係に迫ることはできませんでした。
増大もしくは新生スパインを特異的に標識・操作できる記憶プローブの開発
そこで、我々は、増大・新生したスパインだけを標識・操作するシナプスプローブを作成しました。まず、青色光照射によりスパイン収縮を誘発できるタンパク質PaRac1 (ref 2) に着目し(図1)、この分子が新生・増大したスパインに集積するように分子デザインしました(ASプローブ)。実際にスパインが増大すると、ASプローブが増大スパインに集積することを確認しました(図2)。また、運動学習の1つであるロータロッド学習を負荷したマウスの学習前後でスパインの形態やASプローブの集積を観察したところ、ASプローブは、増大スパインの約70%、新生スパインの80%を標識し、非特異的な標識は2%未満という、感度・特異度ともに高いプローブであることが確認できました(図3)。次に、生きた動物の脳内で、スパインを人為的に操作できるかを確かめました。大脳皮質を広範囲に光照射するため、一次運動野上に光ファイバーを留置したマウスを用い(図4a)、前述のロータロッド運動課題を学習させた後に青色光を照射しました。光照射によりプローブが集積したスパインは収縮しましたが、ASプローブを持たないスパインには変化が見られませんでした(図4)。これは、マウス脳内において増大・新生スパインを特異的に収縮させること、言いかえれば学習・記憶に関連したスパインだけを収縮させることが可能になったことを意味します。
運動学習に伴い増大・新生したスパインを収縮させると、既得学習が失われる
最後に、このような光操作で、マウスの行動にどのような影響が出るのかを調べました。ASプローブを遺伝子導入した群と、していない群に分け、ロータロッド課題を学習させた後、青色光でスパインを収縮させました(図5)。すると、遺伝子導入を受けていない群は光照射の影響を受けなかったのに対して、遺伝子導入群は光照射によって学習成果が低下しました(図5b)。これは、標識したスパインを収縮させることにより既得学習が失われた事を意味します。
おわりに
今回の研究で、増大もしくは新生されたスパインの分布を可視化・操作する方法を世界に先駆けて確立し、スパインの形態可塑性が学習・記憶に必要であることを明らかにしました。スパインは、正常な脳機能だけでなく、心的外傷後ストレス障害や自閉症といった精神神経疾患の病態にも深く関与していることが示唆されています。このような新規シナプスプローブを活用することにより、病態生理がシナプスというミクロなレベルで解明され、新規治療法の細胞基盤としての研究が発展していくことが期待されます。
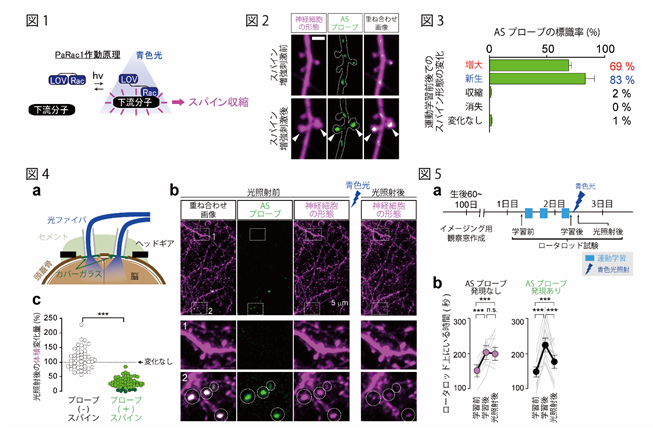
図の説明
図1:PaRac1の模式図。暗所ではPaRac1は下流分子と結合出来ないが、青色光の照射により構造変化をおこし、下流
分子と結合、スパインを収縮させる。
図2:グルタミン酸アンケージング法により白矢印のスパインを人工的に増大させたところ、これらの増大スパインにはAS
プローブが集積した。
図3:運動学習前後でのスパイン形態の変化を5種に分類し、各スパインへのASプローブの集積率を算出。ASプローブ
は、学習後に増大・新生したスパインを約69%・83%の割合で標識する一方、非特異的な標識は2%未満だった。
図4:(a) 観察窓・光ファイバー留置図。両側第一次運動野上に光ファイバーを留置し、広範囲の神経細胞を刺激出来るよ
うにした。(b) 青色光照射前にプローブが集積していたスパイン(白破線枠2)は、照射により収縮する。しかし、プロ
ーブを持たないスパインは影響を受けない(白破線枠1) (c) 光照射に対するスパインの大きさを、ASプローブを
発現するスパイン(緑丸)と発現しないスパイン(白丸)ごとにプロット。青色光照射が、記憶プローブを発現するスパ
インを特異的に収縮させることが分かる。
図5:(a) マウスは両側の一次運動野に記憶プローブを遺伝子導入した群、対照実験として記憶プローブを導入しないマウ
ス群(コントロールプローブ導入マウス)を用意し、どちらの群もロータロッド運動学習後に青色光を照射した。マウス
は、運動学習前、学習後、光照射後の3点でロータロッド運動能力を測定。(b)コントロール群では光照射による影響
は受けないが、記憶プローブを導入したマウス群は、獲得した運動学習記憶を光照射により失うことが示された。
参考文献
1.Matsuzaki M, Ellis-Davies GC, Nemoto T, Miyashita Y, Iino M, Kasai H. Dendritic spine geometry is critical for AMPA receptor expression in hippocampal CA1 pyramidal neurons. Nature neuroscience 2001;4:1086-92.
2.Wu YI, Frey D, Lungu OI, et al. A genetically encoded photoactivatable Rac controls the motility of living cells. Nature 2009;461:104-8.
DOI 10.11481/topics22
掲載日:2016年3月7日 登録日:2016年5月26日
はじめに
脳内の数百億もの神経細胞は、神経突起を介して互いに複雑に連結し合い、膨大な神経回路網を形成しています。これらの神経突起は直接連結しているのではなく、シナプスと呼ばれる数十nmの隙間があります。神経細胞体の興奮がシナプスまで到達すると、シナプス間隙より神経伝達物質が分泌され、放出された伝達物質は隣の神経細胞のシナプス後膜に結合します。このようなシナプス入力が積算されると神経細胞は興奮し、次々と隣の神経細胞に電気的信号を送ることができます。大脳皮質のシナプスの多くは、神経突起上にあるマッシュルームのような小突起(スパイン)の上に形成されます。スパインは、大きさとそのスパイン上に形成されるシナプスの伝達効率が非常に良く相関することが知られています1。重要なことは、スパインが学習・記憶により劇的に増大したり、さらには新しいスパインが構築され既存のスパインが排除されたりする点です。すなわち、スパインの形態変化により神経回路網は動的に変化するので、スパインは学習・記憶・認知の細胞基盤であると考えられてきました。しかし、動物の生きたままの脳内で、増大・新生したスパインを特異的に標識・操作する手法が無かったため、スパインと学習・記憶との因果関係に迫ることはできませんでした。
増大もしくは新生スパインを特異的に標識・操作できる記憶プローブの開発
そこで、我々は、増大・新生したスパインだけを標識・操作するシナプスプローブを作成しました。まず、青色光照射によりスパイン収縮を誘発できるタンパク質PaRac1 (ref 2) に着目し(図1)、この分子が新生・増大したスパインに集積するように分子デザインしました(ASプローブ)。実際にスパインが増大すると、ASプローブが増大スパインに集積することを確認しました(図2)。また、運動学習の1つであるロータロッド学習を負荷したマウスの学習前後でスパインの形態やASプローブの集積を観察したところ、ASプローブは、増大スパインの約70%、新生スパインの80%を標識し、非特異的な標識は2%未満という、感度・特異度ともに高いプローブであることが確認できました(図3)。次に、生きた動物の脳内で、スパインを人為的に操作できるかを確かめました。大脳皮質を広範囲に光照射するため、一次運動野上に光ファイバーを留置したマウスを用い(図4a)、前述のロータロッド運動課題を学習させた後に青色光を照射しました。光照射によりプローブが集積したスパインは収縮しましたが、ASプローブを持たないスパインには変化が見られませんでした(図4)。これは、マウス脳内において増大・新生スパインを特異的に収縮させること、言いかえれば学習・記憶に関連したスパインだけを収縮させることが可能になったことを意味します。
運動学習に伴い増大・新生したスパインを収縮させると、既得学習が失われる
最後に、このような光操作で、マウスの行動にどのような影響が出るのかを調べました。ASプローブを遺伝子導入した群と、していない群に分け、ロータロッド課題を学習させた後、青色光でスパインを収縮させました(図5)。すると、遺伝子導入を受けていない群は光照射の影響を受けなかったのに対して、遺伝子導入群は光照射によって学習成果が低下しました(図5b)。これは、標識したスパインを収縮させることにより既得学習が失われた事を意味します。
おわりに
今回の研究で、増大もしくは新生されたスパインの分布を可視化・操作する方法を世界に先駆けて確立し、スパインの形態可塑性が学習・記憶に必要であることを明らかにしました。スパインは、正常な脳機能だけでなく、心的外傷後ストレス障害や自閉症といった精神神経疾患の病態にも深く関与していることが示唆されています。このような新規シナプスプローブを活用することにより、病態生理がシナプスというミクロなレベルで解明され、新規治療法の細胞基盤としての研究が発展していくことが期待されます。
図の説明
図1:PaRac1の模式図。暗所ではPaRac1は下流分子と結合出来ないが、青色光の照射により構造変化をおこし、下流
分子と結合、スパインを収縮させる。
図2:グルタミン酸アンケージング法により白矢印のスパインを人工的に増大させたところ、これらの増大スパインにはAS
プローブが集積した。
図3:運動学習前後でのスパイン形態の変化を5種に分類し、各スパインへのASプローブの集積率を算出。ASプローブ
は、学習後に増大・新生したスパインを約69%・83%の割合で標識する一方、非特異的な標識は2%未満だった。
図4:(a) 観察窓・光ファイバー留置図。両側第一次運動野上に光ファイバーを留置し、広範囲の神経細胞を刺激出来るよ
うにした。(b) 青色光照射前にプローブが集積していたスパイン(白破線枠2)は、照射により収縮する。しかし、プロ
ーブを持たないスパインは影響を受けない(白破線枠1) (c) 光照射に対するスパインの大きさを、ASプローブを
発現するスパイン(緑丸)と発現しないスパイン(白丸)ごとにプロット。青色光照射が、記憶プローブを発現するスパ
インを特異的に収縮させることが分かる。
図5:(a) マウスは両側の一次運動野に記憶プローブを遺伝子導入した群、対照実験として記憶プローブを導入しないマウ
ス群(コントロールプローブ導入マウス)を用意し、どちらの群もロータロッド運動学習後に青色光を照射した。マウス
は、運動学習前、学習後、光照射後の3点でロータロッド運動能力を測定。(b)コントロール群では光照射による影響
は受けないが、記憶プローブを導入したマウス群は、獲得した運動学習記憶を光照射により失うことが示された。
参考文献
1.Matsuzaki M, Ellis-Davies GC, Nemoto T, Miyashita Y, Iino M, Kasai H. Dendritic spine geometry is critical for AMPA receptor expression in hippocampal CA1 pyramidal neurons. Nature neuroscience 2001;4:1086-92.
2.Wu YI, Frey D, Lungu OI, et al. A genetically encoded photoactivatable Rac controls the motility of living cells. Nature 2009;461:104-8.
原稿募集のお知らせ
一般向け広報活動の一環として、学会ホームページ上に日本神経化学会会員の研究成果を紹介するコーナー「神経化学トピックス」がスタート致しました。
本コンテンツへの掲載をご希望される場合、
自薦、他薦は問いませんので、以下のメールアドレスまで
- タイトル
- 著者
- 掲載誌名
- 簡単な内容説明(論文の抄録でも結構です)
をご連絡下さいますようお願い致します。
候補の中から対象となる研究成果を当会出版・広報委員会で選考し、選ばれた会員の方に執筆要領等をお知らせ申し上げます。
日本神経化学会会員の皆様からの積極的なご応募をお待ちしております。
【ご連絡はこちらまで】
日本神経学会事務局 jsn@imic.or.jp ※@を半角にしてご使用ください。
「神経化学トピックス」に掲載されている内容については無断転載を禁止します。
ただし、大学教員あるいは高校教諭による講義等の教育用に限って自由にご使用下さい。