39. RNA代謝調節因子Ddx20による神経前駆細胞およびオリゴデンドロサイト前駆細胞維持機構
はじめに
脳や脊髄を構成する中枢神経系の発生において、神経前駆細胞の増殖と神経前駆細胞からニューロンやグリア細胞への分化は精密に制御されています。その中でも転写因子は神経前駆細胞の運命決定に重要な役割を果たしており、発生期中枢神経系の各領域における特定のニューロンおよびグリア細胞の産生に深く関与しています。塩基性ヘリックスループへリックス(bHLH)型転写因子のOlig2(Oligodendrocyte transcription factor 2)は、脊髄運動ニューロンとオリゴデンドロサイトの発生に必須の分子であることが知られています1,2。また、Olig2は一部のコリン作動性ニューロンやアストロサイトの分化制御、神経幹細胞の増殖にも寄与することが報告されており、中枢神経系の細胞構築に極めて重要な因子です。しかし、Olig2がこのような多様な機能を発揮するメカニズムについては未だ不明な点が残されています。
新規Olig2結合因子Ddx20は、神経前駆細胞とオリゴデンドロサイト前駆細胞の生存に必須である
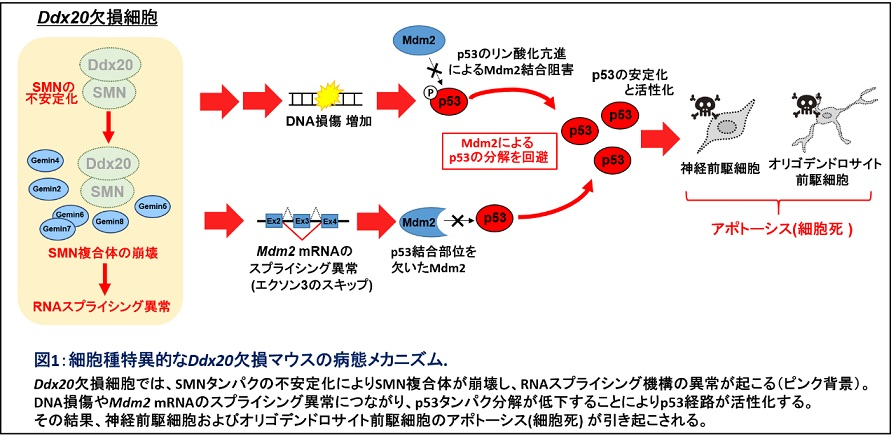
我々は、Olig2に結合する分子によって異なる機能を発揮する可能性を考えて、新規Olig2結合因子を酵母ツーハイブリッド法を用いて探索したところ、RNAヘリカーゼのDdx20(DEAD box protein 20)を同定しました。Ddx20はGemin3またはDP103とも呼ばれ、RNAスプライシング、RNA輸送、マイクロRNA合成、転写調節など多機能な分子として知られています3。中枢神経系発生におけるDdx20の機能を明らかにするために、中枢神経系特異的Ddx20欠損マウス(Nestin-Cre; Ddx20 cKOマウス)を作製し、表現型を解析しました。このマウスは出産直後に死亡し、脳に重篤な奇形が生じていました。詳細な組織解析の結果、Ddx20欠損マウスの脳および脊髄において、神経前駆細胞やオリゴデンドロサイト前駆細胞で細胞死(アポトーシス)が生じており、その過程でp53タンパクが核内に蓄積していました。p53は、さまざまな細胞ストレスにより核内に蓄積し、増殖の停止やアポトーシス誘導に関わることが知られている転写因子です。p53が蓄積する原因を解析したところ、DNA損傷が増加していたほか、p53タンパク分解に関与するMdm2 をコードするmRNAの第3エクソン(Exon3)がスキップしてしまうRNAスプライシング異常も生じていました。Mdm2のExon3はp53結合ドメインをコードしており、このExon3スキップによりMdm2はp53結合能を喪失し、p53の分解が減少することによってp53タンパクの蓄積が起きることが考えられました。Ddx20はRNAスプライシングを制御するSMN複合体の構成因子の一つとしても知られています。興味深いことにDdx20欠損マウスでは、SMNのタンパク量が低下しており、RNAスプライシング機構に異常をきたしていました。これらの結果はDdx20がゲノムの安定化とp53抑制因子Mdm2遺伝子のRNAスプライシング制御という複数の機序でp53経路を抑制し、神経前駆細胞やオリゴデンドロサイト前駆細胞を維持することを示しています4(図1)。
Olig2はDdx20安定化によるRNA代謝制御とp53経路の抑制を介して神経前駆細胞の増殖を促進する
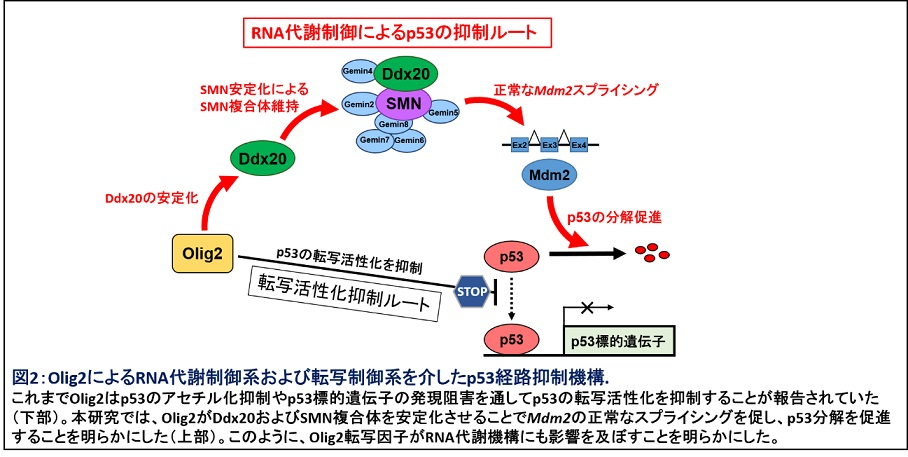
それでは、Olig2とDdx20の相互作用の意義とは何なのでしょうか?私達は、Olig2がDdx20タンパクの安定化に寄与することを見出しました。Olig2を欠損した神経前駆細胞においてDdx20遺伝子発現に有意な変化がない一方で、Ddx20タンパクレベルが有意に低下していました。Olig2欠損神経前駆細胞ではDdx20欠損マウスでも検出されたSMNタンパクの不安定化が生じており、Mdm2のスプライシング異常やスプライソソーム関連分子の異常も認められました。Olig2欠損神経前駆細胞ではp53が安定化し、増殖が抑制されることが報告されています。そこで、Olig2欠損神経前駆細胞にDdx20を強制発現させたところ、p53レベルが低下し、細胞増殖能も改善しました。これまでOlig2がp53による転写活性化を抑制することで神経前駆細胞の増殖維持に寄与することが知られていましたが5、本研究によりOlig2とDdx20との相互作用を介したRNA代謝制御によるp53経路抑制機構の存在が明らかになり(図2)、神経前駆細胞の増殖に寄与する新たなメカニズムを明らかにしました4。
おわりに
本研究によりOlig2による神経前駆細胞の増殖促進メカニズムの一端が明らかになりました。Olig2は転写制御だけではなく、RNA代謝調節にも関与することがわかり、Olig2の多彩な機能が見えてきました。このことはOlig2が様々な細胞の発生・分化に関与する背景の1つになっていると考えられます。一方Ddx20も、RNAスプライシング以外にRNA輸送、翻訳制御、転写調節など多様な機能を有しており、本研究で示したメカニズム以外の機序によっても中枢神経系の様々な発生プロセスを制御していることが予想されます。実際に、Ddx20がオリゴデンドロサイト前駆細胞の維持だけではなく、オリゴデンドロサイトの最終分化の過程にも必須の役割を持つことを明らかにして報告しました6。また、Olig2はグリオーマ(神経膠腫)5やメラノーマ(悪性黒色腫) 7の進行に関わっていることが報告されており、さらに、Ddx20も様々ながんの病態に関わっていることが報告されていますので3、これら2つの因子の相互作用がMdm2-p53経路の制御などを介することで、がんの発生や進行の鍵となっている可能性があります。今後さらに研究を進めることで、先天性神経疾患やがんの病因解明とそれらの治療法開発の糸口になることが期待されます。
参考文献
- Takebayashi H et al. The basic helix-loop-helix factor olig2 is essential for the development of motoneuron and oligodendrocyte lineages. Curr. Biol. 12, 1157-1163, 2002. DOI: 10.1016/s0960-9822(02)00926-0.
- Lu QR et al. Common developmental requirement for Olig function indicates a motor neuron/oligodendrocyte connection. Cell 109, 75-86, 2002. DOI: 10.1016/s0092-8674(02)00678-5.
- Curmi F, Cauchi RJ. The multiple lives of DEAD-box RNA helicase DP103/DDX20/Gemin3. Biochem Soc Trans. 46, 329-341, 2018. DOI: 10.1042/BST20180016.
- Bizen N et al. Ddx20, an Olig2 binding factor, governs the survival of neural and oligodendrocyte progenitor cells via proper Mdm2 splicing and p53 suppression. Cell Death Differ. 2022. Online ahead of print. DOI: 10.1038/s41418-021-00915-8.
- Ligon KL et al. Olig2-regulated lineage-restricted pathway controls replication competence in neural stem cells and malignant glioma. Neuron 53, 503-517, 2007. DOI: 10.1016/j.neuron.2007.01.009.
- *Simankova A, *Bizen N et al. Ddx20, DEAD box helicase 20, is essential for the differentiation of oligodendrocyte and maintenance of myelin gene expression. Glia 69, 2559-2574, 2021. *共同筆頭著者. DOI: 10.1002/glia.24058.
- Lee JE et al. Olig2 regulates p53-mediated apoptosis, migration and invasion of melanoma cells. Sci Rep. 11, 7778, 2021. DOI: 10.1038/s41598-021-87438-x.
このページの目次
「神経化学トピックス」では、神経化学のトピックを一般の方にもわかりやすくご紹介します。
※なお、目次記載の所属は執筆当時の所属となっております。